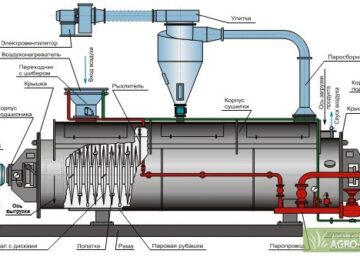
Режим інкубації ікри та температурної адаптації передличинок осетрових риб (Частина 15)
Інкубація ікри. Термічний режим інкубації ікри, отриманої в не традиційні терміни (наприклад, влітку) від плідників осетрових риб, що резервуються при низьких температурах багато в чому визначає ефективність біотехнологічного процесу. Природна температура води на рибоводних ділянках та цехах у цей період значно перевищує регульовану в інкубаційних апаратах цехів за низьких температур. Різниця температур води досягає 10°С і вище. При різкому підвищенні температури води в період інкубації ікри проявляється асинхронність та порушується типовість ембріонального розвитку, що веде до формування таких каліцтв, як асиметрії осьових органів щодо жовткового мішку, викривлення, недорозвинення преанального та хвостового відділів та інше.
Процес вилуплення передличинок при цьому зазвичай дуже розтягнутий. Тому температурний режим інкубації ікри слід програмувати відповідно до планованого терміну вилуплення та розрахункових термінів досягнення різних стадій ембріогенезу. Розроблено три схеми програмованого температурного режиму інкубації ікри в цехах при низьких температурах:
1. Початок інкубації севрюги та осетра здійснюєтьсяз поступовим підвищенням температури дозрівання плідників до природної температури з градієнтом 1,0-1,5ºС на добу. Чим нижча температура води при дозріванні самок, тим триваліший період інкубації. На початку вилуплення передличинок можна прискорити підвищення температури на 2-3ºС за 4 години.
2. Друга схема відрізняється тим, що до 28-ї стадії (прямий подовженої серцевої трубки) інкубація ікри здійснюється за низьких температур води (для російського осетра – 11–13ºС, для севрюги – 15ºС). Подальше підвищення температуриздійснюється з градієнтом 2ºС на добу.
3. Третій підхід полягає в імітації добових коливань температури води: зниження та підвищення в межах 2ºС за добу. При цьому середня температура підтримується протягом наступних двох діб. Це дозволяє керувати терміном інкубації, сповільнюючи розвиток ікри шляхом зниження температури води в інкубаційних апаратах на 3-5ºС. Максимальнадопустима тривалість зниження температури – 4-6 год.
Температурна адаптація передличинок. Вилуплення зародків у цехах за низьких температурах проходить, як правило, при температурах води значно нижче за природну. Тому, для переведення передличинок у відкритий басейновий цех або ставки необхідно здійснювати температурну адаптацію зовнішніх умов.
В цьомуу разі вилуплені предличинки переводяться з накопичувача в басейни або лотки цехів за низьких температур (щільність посадки може бути збільшено до 20 000 – 25 000 тис. шт/м2), підключені до рециркуляційної системи водопостачання.
Через АСУ задається режим підвищення температури з тривалістю 1–1,5 діб до значень температури води у відкритому басейновому цеху та ставках рибоводного заводу.
За даними численних експериментів оцінювалася життєстійкістьпотомства осетрових риб, одержуваного в нетрадиційні терміни приштучне розведення. Необхідність такої оцінки пов’язана з тим, що процес отримання та інкубації ікри проходить при нижчих температурах, ніж подальше підрощування та вирощування молоді.



Адаптація до високої температури довкілля проходить на останніх стадіях зародкового розвитку і в перші дні існування поза оболонкою після вилуплення. Згідно з отриманими результатам, молодь, отримана від севрюги пізнього ходу, за своїм морфофізіологічним показникам нічим не відрізняється від першої молоді половини ходу.
Таким чином, отримання молоді осетрових у нетрадиційні терміни при використанні методу тривалого витримування плідників та адаптації отриманого потомства до температурних умов зовнішнього середовища проходить успішно, що дозволяє суттєво збільшити масштаби промислового відтворення.
Поліфункціональна оцінка фітнес-показників личинок та молоді. Моніторинг якості молоді є важливим елементом штучного відтворення осетрових (наприклад, штучне відтворення на риборозвідних заводах та програми відновлення запасів) і повинно проводитися не лише перед випуском молоді у природні водойми, але і протягом усього технологічного циклу.
Під час моніторингу необхідно здійснювати контроль за відповідністю всіх показників нормативним значенням. Поліфункціональна оцінка необхідна для відбору молоді риб у маткові стада, випуску та товарного вирощування. В останньому випадку молодь повинна мати високі темпи зростання, вгодованість, низькі кормові коефіцієнти і не вимагати жорсткості відбору за адаптивним фітнес-показниками. Ефект доместикації плвдників у маткових стадах на осетрових заводах та отриманого від них потомства, обумовлений штучним відбором на пристосованість до заводських умов і може бути несприятливим для виживанні молоді та стані популяцій у природних умовах.
Крім того, доместикація може призвести до послаблення фітнес показників, що виражається у зниженні опірності до захворювань та екстремальним екологічним впливам, аномаліям відтворювальної системи риб тощо.
Прижиттєві методи оцінки якості та контроль розвитку потомства повинні відповідати наступним основним вимогам:
• включати сукупність показників, які комплексно характеризують функціональний стан вирощуваної личинки та молоді;
• скорочувати час проведення дослідів, травматизм та загибель досліджуваних предличинок, личинок та молоді;
• передбачати можливість оцінки інформації про перспективи подальшого виживання, нормального розвитку, впливу на життєздатність та генетичну структуру популяції осетрових;
• включати систему показників, що екологічно адекватно пов’язані з основними факторами, що визначають виживання молоді після її випуску у природні водойми.
Прижиттєві методи оцінки – експрес-тести. Вказаним вимогам відповідають такі експрес-тести якості потомства осетрових риб, отриманого на осетрових риборозвідних заводах.
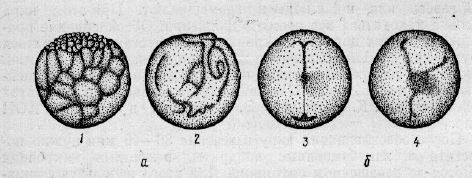
Видоспецифічні особливості реакції предличинок на зміну глибини. Оцінка якості предличинок провадиться з використанням видоспецифічної поведінкової реакції осетрових на перепад глибини. Тільки нормальні життєздатні предличинки можуть здійснювати “Свічки”. Така поведінкова реакція пояснюється природними умовами річки з різним донним покриттям (галькове або мулисто-піщане). На відміну від галькового, дно з мулом менше придатне для предличинок (гірше кисневий режим, велика ймовірність замулювання, наявність дрібних хижаків).
Життєздатні предличинки потрапляючи у несприятливі умови річки, збільшують інтенсивність «свічок», що сприяє їх зносу протягом більш сприятливих ділянок річки. Предличинки з різними морфологічними дефектами головного відділу, серця, жовткового мішка тощо. не здатні, після вилуплення, здійснювати періодичні вертикальні підйоми і в природних умовах річки можуть потрапити в ділянки з більшою глибиною і загинути в внаслідок замулення.
Про якість потомства можна судити за інтенсивності підйомів “свічок”. Інтенсивність свічок у предличинок білуги і російського осетра підвищується в період, наступний за вилупленням.
Після переходу на зяброве дихання частота «свічок» знижується, і предличинки починають здійснювати горизонтальні переміщення, а у момент переходу на змішане харчування ця частота наближається до нуля.
У перші три доби предличинки російського осетра та севрюги найбільш чутливі до перепадів глибин. У білуги реакція на перепад глибин слабша через менше розвинений орган стато-акустики на цій стадії. Відразу після вилуплення проводять тестування з метою оцінки відсотка предличинок, які адекватно реагують на перепад глибини. Цей тест також можна використовувати для оцінки якості плідників за якістю потомства та при відборі личинок для формування або поповнення ремонтно-маткових стад
Плавальна здатність личинки та молоді осетрових риб. Наступним тестом, що дозволяє оцінити життєздатність личинок і молоді осетрових, є тест «реореакція», чи так званий, «реотаксис», який полягає в тому, що, перебуваючи у потоці води риби, як правило, рухаються проти течії.
Даний тест передбачає визначення часу, протягом якого риба може рухатися в потоці води з певною швидкістю. Плавальна здатність молоді осетрових визначається в експериментальних умовах із застосуванням гідролотка з постійною глибиною, аналогічного лотку, починаючи зі стадії вилуплення передличинок. До переходу передличинок на екзогенне харчування, швидкість течії в лотку підтримується рівною 15,8 см/с, а на пізніших стадіях розвитку вона збільшуєтьсядо 20,6 см/сек.
Слід зазначити, що важливе значення у підтримці плавучості та опірності потоку має загальна сформованість тіла та розташування плавників. У перші дні після вилуплення, перед личинки осетрових ще позбавлені плавників, їхній хвостовий відділ слабкий, тому вониз датні здійснювати тільки вертикальні спливання «свічки», здійснюючи їх за рахунок хвиле подібних рухів всього тіла.
З переходом на активне харчування тіло личинок набуває форми, характерної для дорослих риб з великим хвостовим подовженням, особливим будовою рила (рострума), сприяють підтримці плавучості та зменшення опору під час руху. Збільшення часу опірності потоку пов’язане з переходом личинок на зовнішнє харчування.
У період переходу на зовнішнє харчування плавальна здатність личинок білуги становить – 120, осетра – 180, севрюги – 80 сек. (при швидкостітечії – 15,8 см/сек).


Збільшення швидкості течії в лотку до 20 см/сек призводить до зниження плавальній здатності. У личинок севрюги старшого віку плавальна здатність вища, ніж у личинок російського осетра та білуги. Це з особливостями будови тіла севрюги (максимальна товщина тіла на 6,1% менше, ніж у білуги), та її пристосованістю чинити опір потоку води. Так, молодь севрюги, довжиною 22 мм, може чинити опір потоку протягом 48 сек, довжиною 60 та 90 мм – 350 і більше 3600 сек відповідно. У заводський і у «дикої» молоді збільшення плавальної спроможності залежить від довжини риб. Так, молодь севрюги заводського походження, довжиною 45 мм має плавальну здатність – 467 сек, при довжині 77,5 і 128 мм – 1499 і 2536 сек відповідно (при швидкості перебігу в лотку 20 см/сек).
У молоді севрюги, отриманої від природного нересту (при середній довжині тіла 62,6мм), плавальна здатність дорівнює 357 сек, при довжині 68,8, 107,8 та 115мм – 367, 651 та 1390 сек відповідно. Плавальна здатність молоді стерляді також залежить від її розмірів. Так, при довжині 65 мм вона становить 125 сек., за довжини 95 мм – 940 сек і за довжини 125 мм – 1280 сек.
Оцінка розмірів та форми жовткового мішка предличинок. Слід зазначити важливість оцінки розмірів та форми жовткового мішка під час здійснення рибоводно-екологічного моніторингу передличинок, вирощених на осетрових заводах.
Важливим показником деформації жовткового мішка передличинок осетрових є відношення його висоти до довжини (що становить нормувід 0,55 до 0,69). Для деформованого (грушоподібного або подовжено-овального) жовткового мішка дане відношення зменшується до 0,29–0,44. Справді, у разі невеликих розмірів жовтково гомішка (і значної індивідуальної мінливості його морфометричних показників), ендогенні ресурси не забезпечують подальшого зростання та нормальний розвиток на одному з найважливіших етапів – переході до екзогенного харчування. Разом з тим, надмірно великий обсяг жовтка на стадіях диференціювання відділів травної системи також негативно впливає на їх формування, призводячи до затримки секреторної функції епітелію.
(Дивись далі Частину 16)